Cooperation
Humans are a very cooperative species. How did that come about? The question is not easy to answer because of the free riding problem, of which more below. Here we attempt an evolutionary account of the rise and persistence of cooperation.
We start by introducing a basic distinction:
·
An agent who only tries to maximize his expected payoff with no care at all for those of others is self-regarding. He will free ride when possible and generally
shirk from contributing to the group when advantageous to himself; however, he
will engage in mutualism: if cooperating maximizes his payoff, he will
cooperate. For example, if stuck in a
snowdrift with others and wanting to get out, he will shovel if he knows the
others won’t shovel unless he does, but he won’t shovel if he knows others will
shovel anyway.
·
An agent who may or may not care about
one’s own payoff but cares for that of others is other-regarding. He may work
for the benefit of others no matter what, for example by shoveling even if nobody
else does; or, he may punish those who do not cooperate, for example by beating
up those who do not shovel; or he may help shirkers, for example by giving hot
tea to those who do not shovel.
Altruism is a type of other-regarding behavior that occurs when by engaging in it an individual X increases the average fitness of her group, but would increase her own more if she did not engage in it. In other words, X
- confers benefit b on the other n member(s) of her group, so that every other member gets a benefit of b/n-1
- in so doing she pays a cost c herself
- b>c
Note that the motivations behind altruism need not be “nice”. If one punishes selfish free riders at a cost, eventually inducing them to cooperate for fear of retribution, one engages in altruism even if one’s motivation is not the welfare of the group but anger at shirking.
The simplest description of the situation is a classic game, The Prisoners Dilemma. Imagine two players, Joe and Jim. If Joe is an altruist, Jim gets a benefit b while Joe gets nothing and pays a cost c to benefit Jim. The same applies to Jim if the situation is reversed. If both are selfish, nobody gets any benefit or pays any cost. If both are altruists, then each gets b and pays c. The following matrix represents the interaction.
|
Jim |
|||
|
Joe |
|
A |
S |
|
A |
b -c; b -c |
–c; b |
|
|
S |
b; -c |
0; 0 |
|
In the rows and columns of the matrix, S stands for being selfish and A for being an altruist, and in the intersection squares the payoffs on the left of the semicolon are Joe’s and the ones on the right of the semicolon Jim’s. For example, if both Joe and Jim are selfish then both get payoff 0. If Joe is selfish and Jim other-regarding, then Joe’s payoff is b (he gets the benefit of Jim’s altruism without paying any cost) and Jim’s –c (the negative sign represents the fact that c is a cost). Note that S dominates A in that, no matter what Jim does, it’s always advantageous for Joe to be self-regarding, as b > b -c and 0 > -c. In short, selfishness beats altruism in that the payoffs associated with the former are always higher than those associated with the latter.
Let us call reproductive fitness the ability of an entity to produce a copy of itself. Then, if payoffs are ultimately proportional to reproductive fitness most evolutionary dynamics will favor selfishness, resulting in a population made up only of shirkers, each getting a payoff of 0. This can be understood biologically and/or culturally. Biologically, individuals may be hardwired to be selfish or altruists, or to have tendencies towards selfishness or altruism; then, selfish individuals will have more babies with the same tendencies and eventually altruists will disappear. Culturally, the reproductive fitness of a strategy consists in the rate at which individuals adopt it. So, it may happen that individual preferences result in the adoption of strategy S over A, resulting in the spreading of selfishness and the disappearance of altruism.
Of course, it would have been better for every individual if all had been altruists (b-c>0), but this does not change the fact that shirkers have the advantage. And yet, altruism is widespread, from bacteria like Myxococcus xanthus, to multicellular organisms, to human societies. One might reply that although altruists do worse than shirkers within a group, altruists in groups with enough altruists do better than shirkers in groups with many shirkers, and that is why altruists win out. There is a kernel of truth in this. In the 1960’s some claimed that altruistic behavior exists because it is good for the group. However, selection operates primarily at the level of the individual, not the group or the species. Group selection does exist but typically its power is rather limited compared to individual selection.
The answer to our puzzle is that under certain circumstances the ultimate payoff for an altruist is in fact greater than that for a selfish individual, and that such circumstances have occurred in the human case. Before we proceed, we must distinguish two types of behavior producing mechanism:
1.
Proximate,
to be found in human psychology, and ultimately in the human brain: in some
situations our minds work thus and so and certain parts of the brain are
activated leading, or contributing, to certain outcomes. For example, brain scans have shown that in
many humans, especially in males, when punishing transgressors at a cost, areas
of the brain delivering pleasure associated with the performance of a
satisfactory action are activated, and many experimental subjects report
enjoying punishing transgressors.
2.
Remote,
to be found in our evolutionary history, which made a behavior such as
punishing transgressors an advantageous trait.
Note that the two mechanisms are logically independent, although empirically related: the same proximate mechanism is compatible with different evolutionary histories or even with divine design, and the same evolutionary history, at the phenotype level, is compatible with different mechanisms, as different proteins can perform the same biological function. In what follows we shall mainly consider the remote mechanism for the fact that most humans are prosocial namely, they engage in behavior benefitting their group either because they are altruists (they benefit the group but would benefit themselves more by not doing so) or because they are selfish and free riding has a lower payoff than cooperating (they benefit the group because it has higher personal benefit than not doing so).
Behavioral Game Theory
A reasonable starting point is to consider behavioral game theory, which tries to determine how people actually behave in strategic situations by using experimental settings. It turns out that when modeling market processes (e.g., supply and demand situations) with clearly defined contracts the assumption that agents are self-regarding by and large leads to correct predictions. However, in social dilemmas (all gain if most cooperate while each has a personal incentive to defect), or in situations involving fairness, modeling with self-regarding players is not successful. Instead, experimental evidence shows that many players, though not all, engage in strong reciprocity, that is, in
- Altruistic cooperation (cooperating in Prisoners Dilemma situations) but quickly defecting if others defect first
- Altruistic punishment (punishing those who do not cooperate by shirking or being greedy even at one’s net cost as long as such costs are not too high)
To understand the evidence, we need to consider some standard games.
- The Ultimatum game: under anonymity, A and B are shown, say, $10. A is told to offer any amount out of the $10 to B. B may accept or refuse. If B accepts, then the money is distributed accordingly. If B refuses, then nobody gets anything. This is a one-shot game, with the assurance that A and B will not meet again. Real life incarnations of this game are “take it or leave it’ offers such as monopoly pricing of perishable goods or eleventh hour offers.
- The Dictator Game: this is like the Ultimatum game with the proviso that B cannot refuse, so that A may keep all the money, if he so chooses. Real life incarnations of this game occur when one may share a windfall gain.
- The Public Goods Game: Groups of two or more subjects are formed. Each member is given a private account P with points (say 10), redeemable at the end for real money. The game has several rounds, each of which consists in the following. Each player can place points in a common account C or keep them in one’s private account. The players are told that the experimenter will give each 40% of the points in C and that the experimenter will not suffer any economic loss. So, with 4 players, if everyone puts all the points in C, each will end up with 16 points, with a net gain of 6. However, if Joe puts nothing in C while the others put 10 points each, then Joe will end up with 22 points, with a net gain of 12. Hence, if Joe is self-regarding, he’ll contribute nothing. We can see this by using a matrix describing the situation, where:
· in the rows C stands for Joe cooperating (putting his point in the common pot) and D for Joe’s defecting (not contributing to the common pot).
· in the columns C stands for the other 3 cooperating and D for them defecting.
· the numbers in the boxes are Joe’s payoffs.
For example, if both Joe and the others cooperate, Joe ends up with a net gain of 6, which is why the box at the intersection of C and C contains 6.
|
|
C |
D |
|
C |
6 |
-6 |
|
D |
12 |
0 |
One can easily see that Joe’s best self regarding policy is to defect every time: if the others cooperate, by defecting he’ll get more than if he cooperates, and if the others defect he would be a sucker if he cooperated. This is a version of The Prisoners Dilemma. Hence, if everybody is self regarding nobody will cooperate. Real life incarnations of this game often obtain in cases of overuse of shared depletable resources such as water, clean air, land, or fish, or in cases of cooperation ventures in simple societies.
Here is what the experimental evidence shows.
· One-shot anonymous ultimatum games provide evidence for altruistic punishment. Although what’s considered an acceptable offer often varies from society to society, responders punish proposers who have made an offer considered too low by rejecting it. This holds also for high payoffs, comparable to a week’s salary. It turns out that
1. If we consider university students in industrialized societies all over the world, the mean offer is 47%, the modal 50%, SD is 0.05 and rejection rates are very small. In small scale societies mean offers are between 27% and 57% (only 15 such societies have been intensively studied). Still, if all 15 societies are considered, the mean offer is 39%, the modal 37% and SD is 13%.
2. Inter-societal variations seem to be related to different types of modes of production. For example, the Machiguenga (Peru), who traditionally lived in mobile single family units and engaged in so little extra-familial cooperation that they did not have personal names, display high levels of self-regardedness with mean offer of 26%, modal of 25%o and almost non-existent rejection. They also display very little propensity for punishment, as selfish behavior among non-family members is expected.
3. Resource sharing is not a predictor of behavior. For example, the Hadza (Tanzania), are highly egalitarian foragers with extensive meat and honey sharing practices, but have a mean offer of 33%, a modal one of 20%, SD is 0.17 and rejection rate is 25%. In larger camps, offers are generally higher and the rejection rate is 20%. Hadza men hunt alone or in pairs.
Significantly, people accept any offer if it is put forth by a non-person, e.g., a computer program or some random device.
· Third parties witnessing the dictator game often engage in altruistic punishment if the proposer is judged too greedy, as long as the cost is not too high.
· In public good games, most (often about 80% in industrialized societies) cooperate initially but end up by defecting if unable to punish free riders; however, if enough individuals punish defectors, very high levels of cooperation can be obtained, eventually with very little punishment. There are exceptions; for example, the Machiguenga do not cooperate and don’t punish defectors.
In general:
·
Intra-societal variations are not statistically correlated to sex,
wealth, education, or age; their causes are unknown: they could be social,
genetic or a combination of the two.
· Inter-societal variations are statistically related to different types of economic organization and levels of market integration (the rate at which a society engages in market exchanges such as buying, selling, working for wages, etc.). Statistical analysis shows that cooperative practices such as big game or whale hunting and high levels of market integration are positively correlated to high levels of prosociality. There is evidence that framing Public Goods Games situations in terms of familiar activities is significant.
Finally, anthropological evidence from present hunter-gatherer groups strongly suggests that early Homo sapiens groups were egalitarian, at least as far as men were concerned, with no big chief, rather strict monogamy, and a considerable degree of food sharing. However, one need not think that our ancestors were motivated by sentiments of justice for their fellow humans; rather, it is sufficient that humans have a tendency against being dominated (All men try to rule, but if they cannot they settle for equality). In some foraging societies for which there are ethnological records egalitarianism extended to females as well, although it was probably restricted to males.
Not surprisingly, then, most human beings behave prosocially, and some tend to engage in behavior favoring the group even at a net cost to themselves. Some such tendencies are also present, but to a significantly lesser degree, in other primates which also seem to display empathy, reciprocity, a sense of fairness, and a tendency to harmonize relationships. (Whether such displays are associated with the same emotions we feel is a controversial issue; however, evolutionary parsimony, given the close genealogical links between all primates, suggests homology rather than analogy, and therefore similar emotions).
Before we attempt an evolutionary account of human prosociality, we need to get a sense of the time scale involved. Human lineage is only partially worked out, with many points remaining unclear. For example, we don’t know whether Australopithecus afarensis is a true ancestor and how Homo Floresienesis, still alive about 10,000 years ago, is related to us. In addition, many species belonging to our genus Homo existed about which we know little. Still, here is the basic story:
6 million years ago: split from the ancestral line leading to chimpanzees
5.6 million years ago: Ardipithecus kadabba. We have only teeth and a few bones and can say little about it
4.4 million years ago Ardipithecus ramidicus; walked upright for short distances; brain size of 350-400cc.
4 millions years ago: Australopithecus afarensis; about 3 or 4 feet, bipedal (but not as good as we are) and ape sized brain, about 400-500cc. (Lucy is about 3.2 million years old).
3 million years ago: Australopithecus africanus, same size as afarensis but with 500cc brain.
2 million years ago: Homo habilis, an early sophisticated stone tool maker; brain size of 500-600cc.
1.8 million years ago: Possible split between our line and Homo erectus (brain size 850-1100cc.)
1 million/500,000 years ago:
Split between Homo Neanderthalensis (bigger and stronger than we and with bigger cranial capacity--bigger brains, 1200-1900cc.; we have first draft of complete genome) and the lineage leading to Homo sapiens (that’s us). The earliest definite anatomically modern human fossils are from Africa, about 195,000 years old. Molecular clock techniques show that probably our species is about 200,000 years old
More evidence is constantly appearing, e.g., Homo Floresiensis (tools and a 500cc brain with big prefrontal cortex!). It’s important to keep in mind a few things:
·
Whatever the vicissitudes of the genus Homo, we are its only non-extinct species
·
We are genetically very alike, differing
from each other only by about one tenth of one percent of our genes, probably
pointing to a bottleneck
·
The greatest genetic variation is in
southern Africa, which intimates that our species originated there
·
We have existed for a relatively short
time (200,000 years or so)
·
For 95% of our existence we lived in
small bands of hunter gatherers; this intimates that our adaptations are
primarily advantageous for that life.
·
Complex societies arose only with the
domestication of animals and the introduction of agriculture, about 10,000
years ago. Cultural artifacts we take
for granted today were invented a very short time ago; for example, writing is
only about 5,000 years old, and today’s major religions are even younger.
Evolutionary dynamics
To provide an evolutionary account of human prosociality, we need to look at evolutionary processes at a rather abstract level. The main mechanism of evolution is natural selection. Its formal requirements are simple. Imagine a population of individuals with different traits such that
- Individuals replicate and make reasonable copies of themselves, transmitting traits to their progeny; this is called “replication”
- Different traits produce different levels of reproductive fitness in the individuals carrying them; this is called “differential fitness”
Things to which replication and differential fitness apply are replicators. Note that replication and differential fitness are substrate neutral: they occur both with RNA and DNA based organisms. Very likely, they also occur in the case of cultural items such as words (Darwin’s example), ideas, rituals, behaviors, and tunes, in which case they are often called “memes”.
The evolutionary process occurs because differential fitness will favor some traits and disfavor others, thus altering their frequencies in the population. Of course, if no new traits were to appear, the evolutionary process would eventually stop; to keep it going, a third requirement must be satisfied:
·
New traits, arise with sufficient
frequency; this is called “variation”
The simplest case of interesting evolutionary dynamics in one in which replication and differential fitness occur and:
- Variation does not occur
- Replicators confront each other randomly: if the frequency of replicator of type R is, say, 1/4 (25% of replicators are R), then any given replicator has probability ¼ of interacting with an R.
Consider the game of escalation/retreat Chicken, allegedly played by (stupid) teenagers in the 50’s. (A variation of it is played by people, institutions, countries engaged in brinkmanship, or animals when they engage in fighting for mates). Two people drive their cars directly at each other until one (or both) swerves off the road or they crash into each other. Suppose that payoffs are: swerving while other does not: 0; both swerve: 5; neither swerves: -10; continuing on while the other swerves: +10. Here is the matrix, with “S” for swerve and “C” for continue straight:
|
|
S |
C |
|
S |
5,5 |
0,0 |
|
C |
10,10 |
-10,-10 |
Now suppose the following:
·
The traits ‘swerver’
and ‘straighter’ are hardwired
·
The payoffs of the game are proportional
to Darwinian fitness, the number of viable offspring one leaves.
How will the frequencies of swervers and straighters change? Will they reach an equilibrium point? Imagine that a large population is made up of swervers, and suppose we drop a straigther among them. Initially she will do very well because she meets only swervers, and therefore she will leave a lot of straighter offspring. So, the percentage of straighters in the population will increase steeply. However, eventually there will be enough straighters that they’ll meet each other frequently enoughto do worse than swervers. So, if the initial population is composed of both straighters and swervers, there will never be 100% straighters or 100% swervers. Is there an equilibrium point?
The graph representing the system’s evolution is
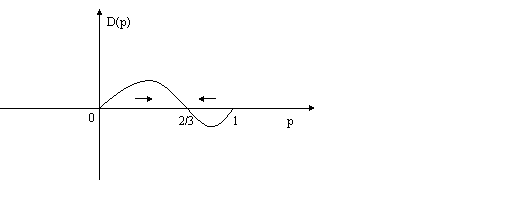
where p is the percentage of swervers and D(p) is the rate of change of p. The system will end up with 2/3 of the players swerving, no matter what the initial distribution is (aside from p=0 or p=1, both of which are, however, unstable). In other words, the system is ergodic. In this system, the equilibrium p= 2/3 is both accessible (the system will get there) and stable (once there the system will stay there).
The previous example is very simple but it gives a clear idea of the evolution of a system by showing the importance of character frequency. A model for the evolution of human cooperation is, of course, much more complex than Chicken; in fact it cannot be handled by analytic means, and therefore one must resort to a computer model. Among the desiderata of a model of the evolution of human cooperation is that it mirror basic human features of hunter-gatherer societies within which we spent 95% of our existence and within which our social evolution took place. A good case can be made that such features are similar to those present in most hunter gatherer societies for which we have anthropological records. Hence, the model should take into account crucial features of late Pleistocene and early Holocene (120,000 BCE to about 10,000 BCE) Homo Sapiens societies including:
- Our tendency to make some errors in the implementation of strategies and in the perception of the situation
- A moderately high discount factor, a quantity measuring the propensity to value future goods as much as present ones
- The fact that groups are small enough (20-30 individuals) to allow direct observation and yet large enough that free riding may be a problem.
- The fact that information about shirking may not be public and accurate
- The fact that interaction is among both kin and non-kin.
- The lack of a central authority capable of enforcing social norms.
- The fact that status differences are limited when compared to agricultural societies.
- The fact that resources are not stored in significant amounts.
- The fact that group membership is fluid in that it typically involves exogamy and high levels of intergroup migration
Finally, the model must describe as outcome an equilibrium that is
- accessible, in the sense that altruism must have a good chance of becoming widespread even if it starts as initially rare
- stable, in the sense that once achieved the equilibrium must be robust.
With this in mind, let us look at some proposed models of the emergence and persistence of cooperation.
POSITIVE ASSORTMENT
MODELS
In standard replicator dynamics, interactions are random. For example, if 20% of the group members are self-regarding, then any given individual will interact with a self-regarding member 20% of the times. However, when it comes to human interaction, such a requirement seems implausible. Hence, many models try to explain the emergence of other regarding behavior by appealing to positive assortment: those with a tendency to be other-regarding interact with each other more frequently than by mere chance. Here are some interesting cases, where c is the cost of conferring a benefit b on others.
Kin altruism
If I increase my identical twin’s fitness at a cost to me, my altruistic genes will be transmitted, through my twin, to the next generation. So, if I behave altruistically towards my kin, we have a case of positive assortment. The key equation here is Hamilton’s rule, which states that kin cooperation is favored by natural selection if the genetic relatedness r (1/2 for siblings, 1/4 for nieces and nephews, and so on) between donor and beneficiary exceeds the cost-benefit ratio of the altruistic act:
r > c/b.
Note that the value of r is unlikely to be high in a population that is not highly inbred. In short, although there is no question that kin altruism is a force in evolution, it cannot explain the simple fact that among primates, and especially humans, altruistic cooperation extends well beyond kin, at times even trumping it.
Reciprocal Altruism
If two individuals are randomly paired to play the Prisoners Dilemma for many rounds, then cooperation becomes probable if a strategy of reciprocal altruism is introduced. The idea here is that X cooperates with Y if Y has cooperated with X, and vice versa. This is an example of positive assortment in that cooperators tend to cooperate with other cooperators more than with the generic player. The simplest of these strategies is Tit-For-Tat (TFT), which says to cooperate if the other player cooperated in the previous round and defect if the other player defected in the previous round: TFT has a short memory. Although the evidence for reciprocal altruism outside humans is relegated to other primates, there is no doubt that it played a role in human interaction, as food sharing in our ancestral past was probably network-based rather than common pot based; in other words, one primarily shared with the individual who had previously shared with one. It turns out that if a few TFT’s are present, TFT beats “Always Defect”, and therefore produces an accessible equilibrium. There is evidence that it is not stable, as TFT is itself supplanted by “Generous TFT” (GTFT), a strategy involving some degree of forgiveness towards shirkers, which becomes stable if it is not too generous. (Of course, what counts as too generous depends on the payoff matrix). However, computer models show that TFT is not accessible and/or not stable when interactions involve more than two players (typically, six players are enough to reduce cooperation drastically) unless there are no mistakes and information is public and accurate, each of which is an unrealistic requirement.
Indirect Reciprocity
Indirect reciprocity is based on reputation. If X benefits Y, then X has a greater chance of being benefited by Z than if he benefits nobody. Positive assortment comes about because those with good reputation will cooperate among each other more than those without it. A strategy embodying indirect reciprocity is the good standing strategy. Players who cooperated with others in the past are in good standing; otherwise they are in bad standing. The strategy is to cooperate only with those who are in good standing if one is in good standing and cooperate unconditionally if one is in bad standing due to a previous mistake so as to reacquire good standing. Under incarnations of this model involving random interaction, indirect reciprocity will succeed if the probability p of knowing the score of another player exceeds the ratio between cost and benefit:
p> c/b.
This information requirement is high, and exceedingly difficult to satisfy if interactions involve more than a few players. Although the use of language may facilitate the attainment of the relevant information, the strategy based equilibrium is unstable if errors are allowed and information is imperfect, as it is bound to be given the incentive to convey false information. There is, however, a way to overcome the information problem in a large group if one engages in costly signaling, namely signaling that cannot be faked. For example, an act of bravery or a public sharing of food or some ritual scarring may increase one’s reputation. So, indirect reciprocity can produce a stable cooperative equilibrium in small groups where everyone knows all the relevant information about everyone else –a fact that can be plausibly modeled with various types of spatial games, or when costly signaling is present.
Networks
In human societies, individuals are part of networks so that they interact only with members to whom they are directly linked. In other words, each individual interacts only with a group of neighbors. After each stage of the game, individuals update their strategy and adopt the strategy C of neighboring cooperators with probability equal to the sum of the payoffs of all the neighboring cooperators divided by the sum of all the payoffs of the neighboring non-cooperators. Nowak and others have shown that cooperation will increase if
1/k > c/b,
where k is the average number of neighbors an individual has. This obtains because the smaller the neighborhoods, the more likely that they are different from each other, thus effecting positive assortment. This means that the smaller the average neighborhood, the greater the chance of an expansion of cooperators. The problem for this model is that in foraging societies the whole group, typically about 30 or so individuals, often constitutes the neighborhood, which requires b to be more than 30 times c, which is unrealistic.
The Bowles and Gintis model
Public goods games, and common experience, indicate that if free-riders are allowed to proliferate, cooperation collapses, eventually degenerating into equivalents of the tragedy of the commons, a typical Prisoners Dilemma situation. The solution to this state of affairs has traditionally been positing some central authority that sanctions defectors, thus insuring cooperation. The reason is simple: since defectors do better than cooperators, how could cooperation evolve unless an enforcer is present? Still, history is replete of societies in which cooperation was achieved and maintained without any central authority because of the propensity of most of their members to engage in altruistic punishment. So, altruistic punishment seems the answer to our question. Gintis and Bowles (2009) have constructed a model that satisfies the conditions set above concerning late Pleistocene human societies and has as an outcome the emergency and persistence of altruistic punishment resulting in widespread prosocial behavior. Here is its rough outline.
Every period, a cooperating individual produces a benefit b, shared by all, at a personal cost c, so that if all cooperate each will obtain b-c>0. Since many other primates display some degree of prosociality, it is reasonable to assume that prosocial behavior in our species is likely to have a biological basis, even if cultural aspects are certainly involved as well. In the model, individuals are haploid (an individual has one copy of each gene) but reproduction is diploid (an individual inherits each gene from one of its parents). Individuals have two relevant loci: one determines whether the individual is selfish or cooperative and the other whether the individual is a punisher or not. Here is how they behave:
- Cooperators always cooperate even if taken advantage of. They have extreme prosocial tendencies and always act on them.
- Selfish types do not cooperate (they shirk) unless sp>c where s is the punishment imposed by punishers for shirking and p is the probability of being punished. p is proportional to the frequency of punishers. In short, selfish types maximize their own expected payoffs. Hence, they do not have prosocial tendencies but act prosocially if it is to their advantage.
- Punishers punish shirkers unless the ratio of selfish to punishers is larger than nmax. Note that this amounts to assuming that punishers have some quorum sensing abilities. Crucially, the cost of punishing a shirker, c is shared by all punishers, so that the more punishers, the less the cost of punishing to each individual punisher. Note that this cost sharing presupposes some degree of coordination. Punishers have prosocial tendencies but act on them conditionally, when it is “sensible” to punish.
- Non-punishers never punish. They lack prosocial tendencies.
Punishers do not punish non-punishers unless they do not cooperate, which means that second-order shirking is not punished. Because of diploid reproduction, four types are possible:
- cooperator-punishers
- selfish-punishers
- cooperator-nonpunishers
- selfish-nonpunishers.
Note that prosocial tendencies and behavior may be complex. For example, cooperator-punishers always act prosocially by cooperating unconditionally, but punish only conditionally. A selfish-punisher punishes and cooperates conditionally. We assume that a selfish-punisher does not punish oneself. All types make mistakes with probability ε; the mistakes arise from bad information about who is shirking, miscalculation of expected payoffs, and execution mishaps. Mutation occurs from selfish to cooperator and vice versa and from punisher to non-punisher and vice versa with probability μ/2. Groups are in a Moore neighborhood with no boundary effects (in the model, groups are on a torus). Individuals migrate with migration rate m and go to a neighboring group; mates are always from a neighboring group. When a group becomes smaller than nmin, it recruits members from neighboring groups. At the end of each period, after punishment has possibly occurred, an individual A is randomly chosen; A reproduces with probability proportional to its payoff relative to the general population’s average payoff by mating with B, randomly chosen from a neighboring group. The single offspring is randomly assigned to A’s or B’s group, and one individual in the population is randomly killed, so that the size of the population is constant. The reproduction procedure just described occurs for a percentage of the population determined by the reproduction rate r. Because of the relation between periods and generations, 25 periods per generation, one period roughly corresponds to one year. The interactions just described account for 10% of any individual’s fitness; the remaining 90% is given by base fitness.
The simulation has the following parameter values
- b=2, c=1, s=2, ε=0.015, nmax=1.7, nmin=4, μ= 0.0001, r= .02
What happens is this. Even if the model starts without punishers, through mutation they appear in a group and initially refrain from punishing because their frequency in the group is too small: they act as sleeping cells, as it were. However, eventually by random drift they become sufficiently frequent in one group to start punishing and therefore compel selfish shirkers to cooperate. This produces two outcomes. First, punishers do not need to punish much, with the result that they do only marginally worse than non-punishers. Second, the average fitness in the group increases so that the group grows in size and since the population is constant it eventually seeds other groups by migration and/or repopulation of undersized groups. Eventually, over 90% of the population is made up of punishers, about 85% or more by selfish, about 15% or less by cooperators, and the rate of shirkers is about 15% or less. These outcomes are stable even if they are not asymptotically stable, as the above mentioned values have small oscillations. Interestingly, some shirkers in the population are necessary for the long term stability of the outcome, otherwise eventually nonpunisher cooperators will take over by random drift, just to be replaced by selfish shirkers. The above results are robust as long as c<b/2 and s>b. Mutation rates do not affect the outcomes much; migration rates up to 50% per generation have little impact.
Since the expansion of altruistic punishers is tied to the expansion of their groups, if absolute resource constraints on group size are implemented (no group can increase in size and intra-group resource competition occurs), no evolution of altruistic punishment or prosocial behavior occurs because by definition prosocial individuals have lower intra-group fitness than some selfish ones. So, if an individual’s chance of reproduction is determined by comparing its fitness not to the average of the general population but to the group’s average, selfishness will win out. However, if 1 indicates absolute resource constraint and 0 none, about 50% of the population is still constituted by punishers even with resource constraints of 0.6, which seems to indicate that under realistic resource constraints altruistic punishment would evolve.
The model presupposes that individuals have the following features:
·
A quorum
sensing ability, especially among punishers. Such capacity is common in
nature even among microorganism, often with spectacular results as is the case
with Vibrio fisheri, the
bacterium responsible for the lighting abilities of some squids. Hence, quorum sensing can be achieved with
simple means; however, presumably in
humans it presupposes sensitivity to the underling psychology of others, an
ability possessed even by small children.
·
An ability
to coordinate action and pass information among punishers. This presupposes some means of communication,
and the obvious candidate is language.
·
An ability
to detect transgressions such as failure to share meat or respect
monogamy. There is evidence that humans
are better at logical thinking when cheater detection is involved than in
identical logical situation of a neutral subject matter.
·
An ability
to punish effectively.
Psychologically this seems achieved by the emotions of anger at the transgression and pleasure at punishing the
transgressor. Physically, this is
achieved through various means, from
withdrawal of cooperation to shunning, ostracism or even violence, which is
especially effective because of our ability in handling sticks and throwing
projectiles.
Note that these requirements may also explain why altruistic punishment is rare in other species.
The model has the following features:
- punishers increase the average fitness of their group. In fact, the increase in fitness arising from being a punisher in a group with generalized cooperation is greater than the increase in fitness in being a nonpunisher in a non-cooperative group. The result is that typically a punisher in a cooperative group will have greater fitness than a random individual in the general population as long as there are enough non-cooperative groups. In general, it is better to be a prosocial individual, even a punisher, in a group with high levels of prosociality than a shirker in a group with high levels of shirking.
- other things being equal, if punishment occurs punishers are less successful than some non-punishers within their own group. For example, selfish-nonpunishers will cooperate when there are enough punishers, which means that they will not be punished, but they will never punish anyone, which means that they will not pay the cost of punishing. In other words, when punishment takes place second order shirkers, those who shirk punishing, will do better than punishers. However, second order shirkers don’t take over because
·
when they reach sufficient frequency
cooperation unravels, and the group’s average fitness, including theirs, collapses: second order shirkers are hoisted by
their own petard
·
once enough punishers are present in a
group, most individuals cooperate without need of punishment, a fact that minimizes
the fitness gap between punishers and second order shirkers.
Multilevel Selection
If human groups compete, their internal organizations become subject to evolutionary pressure. This is a case of cultural evolution in which unsuccessful groups will tend to vanish or change their practices, for example by preemptively adopting some of the practices of more successful groups. The result will be that advantageous group traits will spread. Here we assume that:
·
higher birth rates and higher degrees of
group cohesion resulting from prosocial practices
have been advantageous group traits
·
both birth rates and group cohesion are
measured by average group fitness.
Hence, conflict in which groups with higher average group fitness win out favors the spread of prosocial behavior in the general population.
There is evidence from prehistoric burial sites that inter-group conflict resulting in warfare was common, a state of affairs possibly caused by
o
severe
climate variability during the last 100,000 years, causing
high levels of population displacement.
(So, although a scarcity of resources would have the negative effect of
increasing intra-group fitness competition thus favoring second order shirkers,
it would also propel groups to fight each other for control of territory, thus
favoring the punishers who maintain group cohesion).
o
altruistic
parochialism, the tendency to help members of one’s
own group at a cost, a feature commonly present in humans obviously favoring
groups with it against groups without it.
Sometimes, parochialism extends to hostility towards members of other
groups, races, or religious affiliations.
Parochialism seems motivated by reciprocal expectations, and, like other
types of altruism, is not always fitness enhancing.
Hence, in addition to individual selection based on migration one may assume that a type of group selection took place as well. In short, the evolution of prosociality was probably a multilevel selection affair. Importantly, group selection may overcome part of the added intra-group disadvantage felt by punishers when resource constraints are present. The reason is that resource constrains increase the frequency of shirkers in a group, which in turn diminishes the average fitness of the group, which puts the group at a disadvantage against groups with higher frequencies of punishers.
Note that there are two opposing forces in the model:
(1)
one favoring punishers and resulting
from two components
a. the
fact that active punishers are advantaged with respect to shirkers who are in
groups without active punishers.
b.
the fact that active punishers, by
raising the average fitness in their group, give advantage to their group in
relation to groups without active punishers
(2)
one favoring shirkers, given by the fact
that active punishers are disadvantaged with respect to some shirkers within
their group.
Hence, any factor overall increasing (1) and/or diminishing (2) will hitch a ride on the success of prosociality.
Leveling Factors
Leveling factors are states of affairs that diminish the relative intra-group disadvantage of punishers, thus diminishing (2) above. As a result, some groups will have more punishers, bringing about (1-b). Note that by diminishing (2) leveling factors diminish the effects of resource constraints otherwise resulting in greater intra-group competition. The overall outcome is that leveling factors
·
increase the speed at which prosociality evolves
·
allow prosociality
to evolve when resource constraints are strong.
The leveling of intra-group fitness differences can come about through various conventional cultural institutions such as monogamy, meat-food sharing, consensual decision-making, distaste at being dominated, all observed in foraging societies. These are conventions, that is, Nash equilibriums, and therefore best replies to themselves, which entails that even selfish individuals will follow them. Further, some type of selective assortment or segmentation may occur, for example, by prosocial individuals dealing more among themselves than with shirkers or even by prosocial individuals expelling shirkers.
Morality
Two aspects of
morality
Morality has two related aspects:
1.
Engaging in prosocial
behavior
2. Thinking
of behavior in terms of motivating moral judgments, namely judgments involving
concepts such as obligation, desert, or punishment. Let us call this moral thinking.
Note that
·
(1) does not presuppose (2); for
example, bees are very prosocial, but they don't
engage in moral thinking. Whether some
apes engage in moral thinking is a complex issue we do not address here. What is very likely is that other members of
the genus Homo (e.g., Neanderthalensis
or Floresiensis)
did. Keep in mind that we are just one
of the many species of the genus Homo.
·
(2) probably presupposes (1) or at least
some degree of prosocial tendencies to generate or
reinforce the motivating strength of moral judgments
·
(2) says close to nothing about the
material contents of moral thinking.
Different groups may systematically reach different moral conclusions
about the same state of affairs even under ideal circumstances.
·
(1) and (2) are associated with emotions
such as anger, shame, and guilt.
·
Evolution has not made us “savages” and
morality is not just a thin veneer over our “animal” nature. Civilization is not a fight against
evolution. On the contrary, nature has
made us so that we engage in prosocial behavior
within our group. By and large, our
deficiencies are not so much in prosociality, but in
excessive parochialism.
When thinking about an evolutionary account of moral thinking several possibilities come to mind. Morality could be
1.
An adaptation increasing the fitness of prosocial individuals in groups.
2.
An adaptation increasing the average
fitness within the group
3.
A neutral trait, like hair color.
4.
A byproduct of an adaptive trait.
Some evidence for the view that (1)-(2) is a good bet is given by the fact that moral thinking
·
works as a very powerful leveling factor within the group by restraining some
shirking, including second order shirking
·
reinforces strong reciprocity in groups
by providing a socially accepted justification for punishment
So, moral thinking
increases intra-group leveling and
therefore piggybacks on prosociality. Of course,
the moral thinking of a group involves
substantial amounts of socialization of the young transforming external
norms into internal preferences. The
result is that morality probably results from a combination of genetic
predispositions and cultural preferences.
Some metaethical considerations
Not all evolutionary models of morality need reach the same conclusions, and therefore what model one adopts matters. For example, if our model had ended up with a vast majority of cooperator-punishers, one could argue that in most of us not only engage in prosocial behavior but also have marked prosocial tendencies. Most of us would then be psychological altruists, benefitting others for their own sake, and this would provide some support for the innateness of morality or for a developmental vindication of morality. However, in the present model most of us are selfish-punishers, with limited prosocial tendencies, and the key to the model’s operation is the role of punishers. The model does not contradict the fact that we have a tendency to engage in comforting behavior; rather, it maintains that selfish impulses are very strong.
Hence, if the previous account of the genealogy of moral thinking is roughly correct, then the most natural, but by no means inevitable, conclusion to reach is that moral thinking tracks the reactions that punishers in one’s group would have to one’s actions. Information on how punishers would behave rests on how they have previously reacted to similar actions. In fact, we are “mindreaders”: we have the ability to attribute mental states to others and predict their behavior from an early age. Of course, the tracking need not be exact as long as it is sufficient to favor prosociality.
If one makes the reasonable assumption that a belief in the intersubjectivity of morality increases its functionality, we should expect that most humans view morality as intersubjective, binding within the group, and possibly, but not necessarily, as universally binding. So, at a minimum, a statement like “Not sharing is wrong” would have a different status from expressions of subjective preference such as “I like berries”. In addition, we should expect that moral rules would have a particular authority so that their violation is especially serious and few, if any, individuals may release us from them, in contrast with rules of etiquette, for example. These expectations are in fact borne out as not only adults but also children as young as three make the moral/conventional distinction in terms of seriousness and authority independence, and generalizabilty. (Young children seem to adopt a radically objectivist view according to which morality is not relative to any group of subjects, and a similar position seems part of commonsense metaethics). Note that such intersubjectivity and authority could be achieved either by believing that moral statements are true, or by believing that the emotive reactions to them are natural and therefore proper, or simply because moral judgments are associated with certain emotions.
In addition,
- Almost all moral systems at some time or other, and most at present, should share a core favoring prosociality, and therefore they should contain injunctions against harming the group and in favor of beneficence towards group members and, almost certainly, some injunctions favoring parochialism. Of course, how different moral systems achieve no-harm and beneficence policies varies in relation to different situations, non-moral beliefs, and historical contingencies.
- There should be moral injunctions geared to publicly reinforcing group cohesion, and therefore practices embodying costly signaling such as ritual scarring, notable resource giving, or participation in resource intensive ceremonies.
- One would expect some moral injunctions that are just fitness neutral or even fitness diminishing conventions, namely accidentally historically determined Nash equilibriums. The persistence of fitness diminishing practices is explained by cultural preferences.
So, while norms against harm and in favor of beneficence and group cohesion, including parochialism, should be almost ubiquitous, remaining moral norms would vary across cultures quite dramatically. In particular, one would expect to find the greatest moral divergence with regard to types of behavior not causally linked to group fitness such as how to treat household animals, or animals not related to the food supply, body ornamentation, or funerary practices.
Evolution tells us that moral thinking was adaptive. Let us make the plausible added assumption that most of it still is. Does all of this vindicate moral thinking, namely provide a reason why one should follow group rules?
There are four non-mutually exclusive main reasons for following social rules:
1. Instrumental:
following social rules is conducive to the welfare of group members; not
following social rules is not.
If morality is an adaptive trait, then it is useful in terms of reproductive fitness, and one can argue that typically reproductive fitness is positively correlated to individual welfare. Of course, a population explosion may lead to a decline of individual welfare, but we can separate welfare from amount of progeny and still favor the rules leading to individual welfare. One might distinguish egoist from altruist instrumental reasons.
·
Egoist:
i.
my following rules reinforces
cooperation, which increases my welfare
ii.
my transgression may be punished, thus
decreasing my welfare.
·
Altruist: my following rules is good for
other group members
Note that egoist and altruist reasons need not be incompatible and the egoist instrumental justification is impotent against Gyges ring scenarios.
2. Opinion based:
following social rules puts one in good standing in the eyes of the group and
one cares about that.
Evolution has made us so that we care about the opinion others have of us and following rules gives a good reputation. One may challenge the reasonableness of caring about what others think of us, but one may counter that good standing avoids punishment, is important in reciprocal relations and more generally for one’s participation in society. Note that this collapses opinion based into egoist instrumental reasons that are impotent against Gyges ring scenarios.
3. Developmental:
following social rules leads to one’s flourishing because they agree with one’s
nature.
One may argue that evolution has made us prosocial, and therefore breaking group rules grates against our nature and renders us unhappy. However, the model does not require that all of us be fully prosocial by nature (have prosocial tendencies), as cooperator-punishers are, but only that most of us behave prosocially; in fact, the model ends up with a majority of selfish punishers. To be sure, humans have some degree of innate altruistic motivation, as evidenced in children’s comforting behavior, but public goods games show that selfish motivation are very powerful. In short, with respect to prosociality there isn’t such a thing as human nature. Hence, the developmental justification applies only to some.
However, to the extent that socialization has transformed norms into wants by internalizing them, a properly socialized individual has a reason for following social rules because doing otherwise will make him feel guilty.
4. Epistemic:
the judgments underlying social rules are true or probably true.
Humans view morality as intersubjective and there is some evidence that objectivity is the default common sense metaethical position. However, whether moral judgments like “Not sharing is wrong” are true or not is a different issue, depending in part on what moral facts are.
·
If one adopts moral realism, the view that moral facts are constitutively
independent of human opinion, then it is likely that moral judgments are not
true. The falsity of moral statements
need not affect the functional aspects of morality; while perceptual faculties
must track truth to be evolutionarily advantageous, moral faculties need not: a
belief in the spirit of the forest may produce as much ecologically sound
behavior as the latest ecological theory.
Of course these considerations do not
prove that moral statements are false or even lack truth value (they are
neither true nor false, much like commands or emotional outbursts like “Booo!”); it is possible that we have moral cognitive
abilities that track moral truths.
However, the genealogy of moral thinking puts the burden
of proof on the supporters of the position that moral statements are true, and
the presence of systematic moral disagreement even under ideal conditions makes comparisons with the natural
sciences suspicious.
·
If one rejects moral realism and adopts
a variety of constructivism to the
effect that moral facts are just facts about possible societal reactions to
one’s actions, a natural conclusion from the evolutionary account of morality
just considered, then one can argue that morality tracks such facts. So, to the extent that one is satisfied by
judgment, emotion, or response dependent accounts of moral thinking, one may
claim that an evolutionary account supports the idea that moral judgments are
true, and this vindicates moral thinking.
Note that the above possible reasons for following social rules are unlikely to apply to the whole of morality. For example, fitness lowering customs or accidentally historically determined ones are likely to be supported, if at all, only by our desire to be part of a group and be well regarded by other members.
An evolutionary
account of religion?
It may be possible to attempt an evolutionary account of religion as well. This is an enormously complex task, involving, among other things, little understood areas like late Paleolitic (alleged) religion, and new fields like cultural evolution and the neurology of religion. Hence, what follows is speculative.
One can speculate that the evolutionary cause for the existence of religion is tied to the functional role of religion in the reinforcement of morality, whose role, in turn, is to reinforce prosocial behavior. In short, human religion exists because of human prosocial and moral behavior, not the other way around. This presupposes that successful religions, and therefore theistic based religions (although religion need not be theistic, presumably, most theistic beliefs have been associated with religions) share a moral core conducive to a type of in-group morality favorable to prosociality. Typically theistic based religions preach
o
helpful attitudes towards correligionaries
o
some sort of supernatural punishment for
transgressors
These are leveling factors favoring prosocial behavior. In addition,
o
religious parochialism
o
group-supporting practices such as
meetings and rituals
favor group cohesion by providing psychological support and costly signaling (e.g., scarring, time-consuming rituals, resource-intensive religious practices).
Presumably, the evolutionary advantage of religion became very significant once humans started living in large groups such as large bands or tribes (in the high 100’s) containing significant inequalities together with an increase in the production of offspring. In particular, the neolithic domestication of crops and animals about 10,000 BCE coincided with the introduction of even larger sedentary communities (in the 1000’s) characterized by theocratic government that justified political authority, unequal distribution of wealth, and parochialism.